Storytelling comes naturally to humans, but since we live in an unnatural world, we sometimes need a little help doing what we’d naturally do.
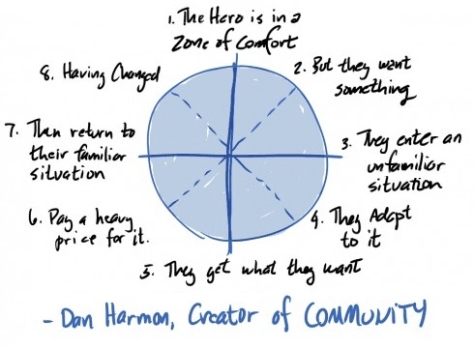
I’m a big fan of the work of Dan Harmon, writer of amazing tv shows like Community and Rick & Morty, and I’ve often heard him talk on his podcast (‘Harmontown’) about his ‘story circle’: a pattern to which most good stories conform.
The REAL structure of any good story is simply circular – a descent into the unknown and eventual return – and that any specific descriptions of that process are specific to you and your story.
The structure itself is pretty simple: a circle, divided and numbered as below, with each number representing a step on our journey.
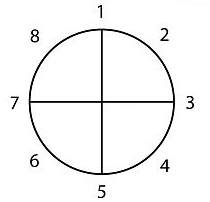
- When you
- have a need,
- you go somewhere,
- search for it,
- find it,
- take it,
- then return
- and change things.
It got me thinking about whether research papers fit this kind of pattern, whether they should, and also whether thinking about such a pattern when developing our papers would help structure them better.
“But my paper is about SERIOUS RESEARCH,” you might say. That may be so, but don’t you want your readers to also enjoy reading about it? If your paper isn’t compelling, then will it even leave a lasting impression?
I’ve taken most of this from Harmon’s notes on the Channel 101 page on story structure, and it’s worth going through that to see some more examples of how some of our favourite movies (ok, he mostly focuses on Die Hard) fit the pattern.
Let’s go through the steps, and see how this structure might relate to the typical academic paper.
1. Establish a protagonist
Who is the protagonist in a research paper? This is – on the face of it – a tricky question, particularly as we are often taught to write in a weird passive (third-person?) voice, resulting in bizarrely disjointed sentences. The crusade against such an impersonal style includes things like the ‘by zombies’ meme, but still some people push it for reasons like ‘the focus should be on the results’, or ‘science should be impartial’. I agree, in that your research should not be carried out with an agenda. But you came up with the question, you designed the experiment, you carried it out, and you are presenting and interpreting the results. You are taking the reader on a journey with you. Don’t write yourself out of it.

In fact, the context of a research paper means your reader instinctively knows who the protagonist is: a scientist, pluckily trying to advance their field. You don’t need to flesh it out any more than that. But you do have to establish the ‘zone of comfort’: the current state of research in your field. By bringing others into your zone at the beginning of the story, this is how your reader identifies with you.
2. Something ain’t quite right
Things aren’t perfect. They could be better.
Science in a nutshell. We could always understand things better than we currently do. Your job here is to show the reader what’s missing, what the gap in your knowledge is. This is your call to adventure.
3. Crossing the threshold
You are now entering an unfamiliar situation, because that’s what science is about: driving into uncharted territory, in search of something more. Harmon says here to figure out what your ‘movie poster’ is; that maybe doesn’t work as an analogy for scientists, as we have a tendency to front-load presentations of our research with the results. The threshold we are crossing here is between defining our knowledge gap, and attempting to rectify that gap with further research. Our movie poster is not the results, but the question.
4. The road of trials
Is there a more fitting descriptor for a methods section? Harmon talks here of how, in ‘Hero with a Thousand Faces’, Joseph Campbell “evokes the image of a digestive tract, breaking the hero down, divesting him of neuroses, stripping him of fear and desire”. You’ve crossed the threshold, the adventure has begun; “our protagonist has been thrown into the water and now it’s sink or swim”.
Your reader needs to know the tools with which you are going to approach your call to adventure, and they need to know this as quickly and efficiently as possible.
The purpose here has become refreshingly – and frighteningly – simple.
5. Meeting with the goddess
This is a time for major revelations, and total vulnerability
…In other words, the results of your research. What did you find? Don’t be afraid to say, we found nothing – particularly if your trials were designed well enough that you can say that conclusively.
To paraphrase Harmon’s words here: we started from a position of safety and comfort (1), but a lack of completion (2) drew us to a question (3) and we were pulled across a threshold into the unknown. Via our experiment, our understanding was transformed (4) by gaining this new, hitherto-unknown knowledge (5). Show it with a definable moment, advises Harmon. Clear, understandable figures and tables bring the reader with you in these revelations.
On the circle, this is the opposite of the protagonist’s zone of comfort; movement beyond this point requires you to push forward. Figuring out the question, and performing the experiment, was almost the easy part compared to what comes next…
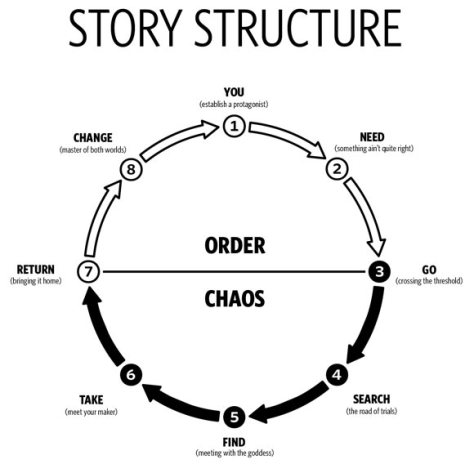
6. Meet your maker
This half of the circle has its own road of trials – the road back up. The one down prepares you for the bed of the goddess and the one up prepares you to rejoin the ordinary world.
I like the description Harmon gives in a later page:
The hardest part (both for the characters and for anyone trying to describe it). On one hand, the price of the journey. The shark eats the boat. Jesus is crucified. The nice old man has a stroke. On the other hand, a goal achieved that we never even knew we had. The shark now has an oxygen tank in his mouth. Jesus is dead- oh, I get it, flesh doesn’t matter. The nice old man had a stroke, but before he died, he wanted you to take this belt buckle. Now go win that rodeo.
With the results in hand, you now have to do the hardest part: interpret what they mean. The ‘heavy price’ we pay as scientists is that sometimes we don’t find what we expect, and things are often messy, and complicated, and hard, and we have to think about them a lot. Again, look at the opposite side of the circle, which was the call to adventure (2). You identified a knowledge gap, and now you need to consider what your results mean in that context – and bring your readers with you.
Now, instead of reacting to the forces of the current state of research – “adapting, changing, seeking” – you have BECOME the current state of research. You are the cutting edge.
You have become that which makes things happen. You have become a living God.
(So I guess this is about where you start fantasising about submitting to Nature)
7. Bringing it home
It’s not a journey if you never come back.
The return to the familiar situation – we can now come back to the beginning of the story, the state of research as we left it, armed with the new knowledge gleaned on our journey.
8. Master of both worlds
The protagonist, on whatever scale, is now a world-altering ninja. They have been to the strange place, they have adapted to it, they have discovered true power and now they are back where they started, forever changed and forever capable of creating change. In a love story, they are able to love. In a Kung Fu story, they’re able to Kung all of the Fu. In a slasher film, they can now slash the slasher.
…And in a science paper, you can now science the science. How are you capable of all this? Because of what happened before – just look at the opposite side of the circle. The opposite of (8) is (4), the road of trials. You did a science, and now you can science some more. Show your reader what you’ve done. The state of research in your field is forever changed, because of your journey. A hero’s journey.
To finish, Harmon provides an example of breaking down the structure using The Matrix: the story of an everyday guy (1) that gets a weird call (2) and, upon following it, realizes that reality was an illusion (3). He learns the ropes (4), talks to the oracle (5), loses his mentor (6), goes back (7) and saves the fucking day (8). Can we apply this kind of structure to our favourite science papers?
Andersson (1982) Nature
Malte Andersson believed Darwin’s hypothesis about the evolution of male sexual ornaments through female preferences was plausible (1), but saw little experimental evidence that such preferences exist (2). Would experimental increase of ornaments confer higher mating success (3)? He manipulated widowbird tail length (4), found males with elongated tails won more mates than those with shortened or ‘control’ tails (5), and excluded other variables to conclude that changes in tail length caused the differences in attraction (6). His results supported Darwin’s hypothesis that certain male ornaments are favoured by female mate choice (7), and probably evolved through it (8).
Not as snappy as the Matrix, sure – but Andersson’s journey changed the idea of sexual selection as ‘plausible with no great evidence’ to ‘plausible with some solid evidence’. A basis for further journeys. For more heroes to come.
Will knowing this structure make you a great writer? No. To return to a quote from earlier:
The REAL structure of any good story is simply circular – a descent into the unknown and eventual return – and that any specific descriptions of that process are specific to you and your story.
You still need a story to tell. But this might just help you figure out how to tell it.
——————-
Quotes all taken from Harmon’s ‘Story Structure‘ posts on the Channel 101 wiki.
——————-
Update: Through the delights of twitter, I just found that Ian Lunt has a video online of a talk he gave on using this approach for science communication, called ‘Shaping stories to save the world‘. I highly recommend watching this, particularly as Ian discusses the difference between the circle and the ‘hourglass’ approach to telling a story. It comes to mind that the story circle may be better suited to seminars and popular writing than it is to research papers – something to think more about, anyway…