
In my last post, I gave a quick overview of my week curating the @realscientists twitter feed, but focused – unsurprisingly – on the delights of the ‘humpoff’ and its ensuring press coverage! As a new scientist takes the reins of that account each week, condemning the previous tweets to the depths of the internet, I thought I would collect my science-based tweets into a post here. Mainly to show how my week was basically an exercise in drawing diagonal lines. I’ll begin here with my tweets from the first day of my week in charge, lightly edited to add a mild bit of coherence:
Let’s start with a little background… obviously, no week on evolution would be complete without some Darwin quotes (taken from ‘Darwin and Genetics‘, an open-access paper by Brian Charlesworth & Deborah Charlesworth):
“The power of Selection, whether exercised by man or brought into play under nature through the struggle for existence and the consequent survival of the fittest, absolutely depends on the variability of organic beings. Without variability, nothing can be effected; slight individual differences, however, suffice for the work, and are probably the chief or sole means in the production of new species.” (DARWIN, 1868)
CHARLES Darwin was the first person to appreciate clearly that evolution depends on the existence of heritable variability within a species to generate the differences between ancestral and descendant populations. The development of Darwin’s thoughts on the nature and causes of evolution is clearly documented in his “transmutation” notebooks of 1836–1838 (BARRETT et al. 1987). Once he had decided that species originated by “descent with modification,” Darwin quickly realized the need to find a mechanism for accomplishing the changes involved. In formulating the idea of natural selection, he was greatly influenced by the experience of breeders in artificially selecting populations of domestic animals and plants. Chapter 1 of The Origin of Species (DARWIN 1859) is famously devoted to documenting the existence of variability in these populations and the effectiveness of artificial selection:
The key is man’s power of cumulative selection: nature gives successive variations; man adds them up in certain directions useful to himself” (DARWIN 1859, p. 30).
It was only a short step to applying this observation to selection in nature:
Can it, then, be thought improbable, seeing that variations useful to man have undoubtedly occurred, that other variations useful in some way to each being in the great and complex battle of life, should sometimes occur in the course of thousands of generations? … This preservation of favourable variations and the rejection of injurious variations, I call Natural Selection (DARWIN 1859, pp. 81–82).
In essence, Darwin identified three conditions that are necessary for evolution via natural selection. First off: a population must exhibit phenotypic variation. ‘Phenotype’ is a fancy word for the set of observable characteristics of an individual. An individual’s phenotype can be anything we measure: body size, shape, development time, lifespan… (And, yes, their behaviour too! But let’s leave that for a little bit later). Have a look at people around you: you (hopefully) all have the same number of fingers and toes, but what about something like, say, height? If you measured the heights of a population (and grouped number of ppl by height), your graph might look like this:
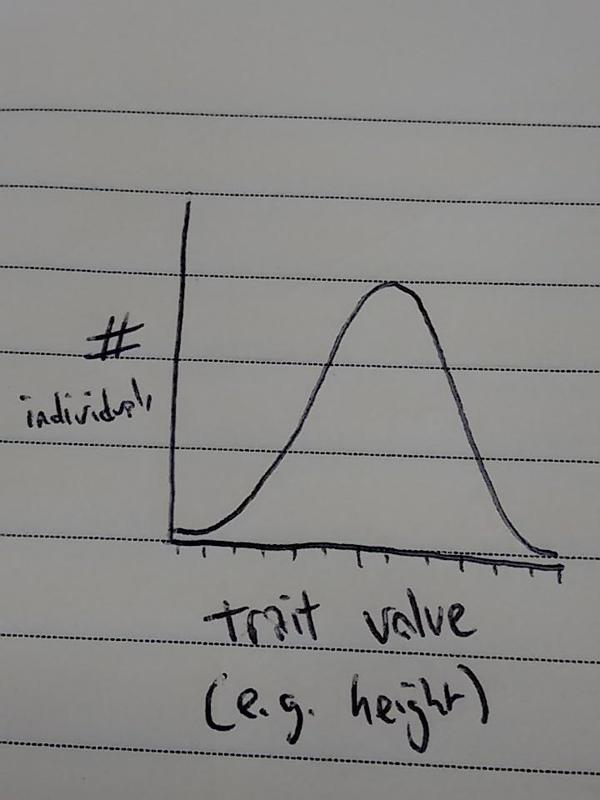
So: within a given population, individuals must be different from each other. What next?
Darwin realised that (ii) these variations must be associated with differences in survival and reproduction. (Hmm… as a very short man, I now regret using ‘height’ as the example trait in my previous tweet… ). Anyway! Individuals must be different from each other, and these differences create variation in how they survive and reproduce…
(iii) Variation in traits must be heritable: parents and offspring must resemble each other in these traits.
Famously, Darwin was unaware of Mendel’s work, meaning that he did not know what the mechanism of inheritance was. On Wed, I’ll talk a little abt quantitative genetics, which helps us understand the inheritance of traits that vary in a continuous fashion (NOTE: I don’t think I did). For now, let’s consider what contributes to phenotypic variation: obviously genes, but what else?
An individual’s environment can strongly affect its phenotype. Recently, the UK govt has come under fire for considering dropping free school meals – health professionals and academic studies claim that diet quality affects academic performance: environment changes the phenotype! Broadly speaking, that would represent a change in the population mean due to environmental effects: i.e., as diet quality (envt value) increases, so academic performance increases.
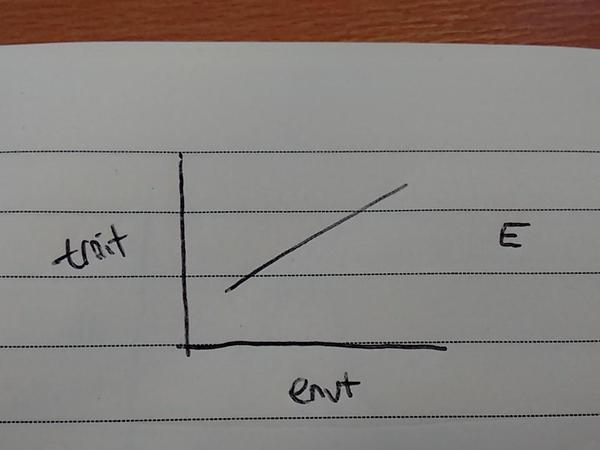
But we also know that different genotypes vary in trait value:

…and genotypes can differ in how they respond to environmental change: known as genotype-by-environment interactions:
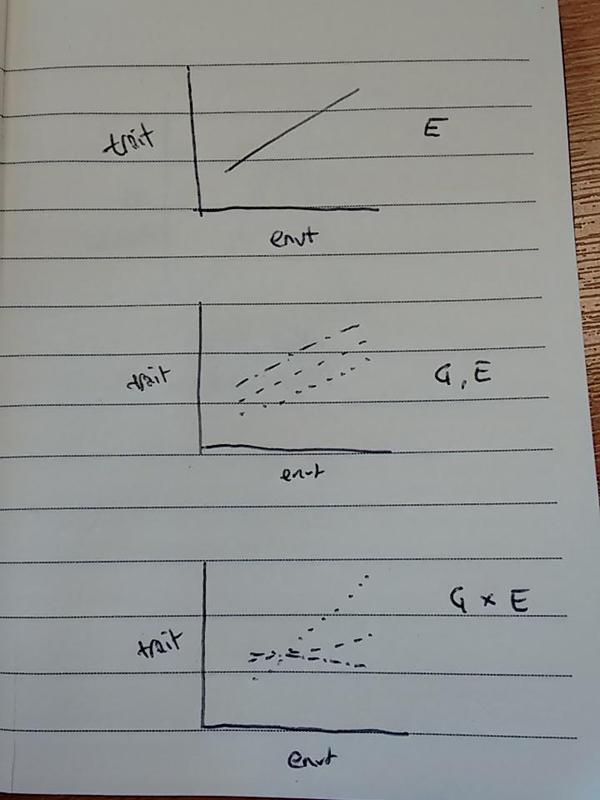
So: heritable phenotypic variation is needed for evolution by natural selection, and phenotypic variation can be affected by G, E, GxE! Excitingly, environmental effects can also change the extent of the variation available for selection to act on:

These lines represent what we call ‘reaction norms’: a reaction norm is a function that describes the change in a genotype’s phenotype over a range of environments. The ability of a genotype to express different phenotypes as environmental conditions change is known as ‘phenotypic plasticity’. Plasticity enables an organism to ‘fit’ its phenotype to the changing environment. Let’s look at a couple of examples…
The desert locust Schistocerca gregaria has a solitary form, in which individuals are well camouflaged and avoid others. However, in their gregarious form they are more active, brightly coloured and form vast migrating swarms.

This change is elicited by repeated physical contact, e.g. through touching or jostling, producing the plastic response. Rogers et al tickled locust legs repeatedly w/ paintbrushes, causing full behavioural ‘gregarisation’ within hours!

You can watch a rather melodramatic video about this experiment:
https://www.youtube.com/watch?v=uURqcI08IC4
It’s since been found that the mechanism is serotonin-related – read more about this here.

This is cool, but I’m more interested in plasticity in ‘labile’ traits: those that are expressed repeatedly across an individual’s lifetime. For example: song! Plenty of animals sing to mark out territories or attract mates, and this is a trait that is expressed repeatedly. There has been a lot of interest in how animals can respond / have responded to the increase in background noise caused by human activity (@c_n_anderson then sent me two reviews on this topic: Patricelli & Blickley 2006; Barber et al 2010). In one experiment, Verzijden et al recorded territorial male chiffchaffs singing along quiet riverbeds and near busy highways. They then raised the background noise at riverbed territories to highway levels, and recorded those birds again… and, finally, recorded the same birds again at normal riverbed background noise (having previously increased it). Here, you can see the plastic response of the riverside birds in minimum song frequency as background is manipulated:

So, why aren’t all animals all plastic all the time? There are costs and limits to plasticity; one such cost is ‘environmental mismatching’. What if there’s insufficient environmental information, or the wrong environmental information is used? The phenotype is then not the ‘right’ one for the environment. This is a particular problem for ‘developmental plasticity’, where individuals use information about their environment to ‘predict’ the optimal phenotype. There’s a nice paper by @mkasumovic on ‘socially cued anticipatory plasticity’: Kasumovic & Brooks 2011. So, that might be bad when the trait is fixed at some point, but what abt individuals that can change the trait in a flexible fashion?
Firstly, let’s keep in mind that assessing the environment and changing trait values is likely to be costly (in time and resources – we’ll talk more later about the importance of resource costs). But also, plasticity itself – the ability to change – may be under selection! Not just the intercept, but the slope of the reaction norm! Plasticity can be adaptive (producing a phenotype in the same direction as the optimal value in the new environment), but can also be maladaptive. Only if the plasticity itself can be shown to have been moulded during evolutionary history to be more effective than a canalised (fixed) phenotype can we consider it to be adaptive. We’ll talk a little more about this tomorrow, but here’s a taster: Earlier, we considered a bird that changes its song frequency depending on background noise. What if only some individuals did that, while others maintained a steady frequency, no matter what the background noise. In some environments, plasticity is undoubtedly better. BUT… what if, in general, this actually gave no survival / reproduction advantage over time? In that case, while the ability to modulate frequency might appear to be adaptive, there’s no evidence that this is the case! MIND. BLOWN.
Oh, and also let’s remember selection acts on ‘extended phenotypes’. Fig from Bailey (2012):

(…but feel free to dust off your copy of @RichardDawkins‘ excellent book as well!)
This resulted in a brief digression about studying plasticity in extended phenotypes using spider webs, and to social behaviour among spiders:
Michelle LaRue then tweeted me her recent paper, which posited that philopatry in king penguins was life history plasticity – perhaps an important way for animals to deal with climate change. This hints at some reasons as to *why* we are interested in plasticity: it lends to the idea of populations moving to novel environments, or coping with changes in environmental conditions.
And, with the sounding of a CLIMATE CHANGE KLAXON, my first day at @realscientists came to an end!

